Hücre Zarı (Plazmalemma)
Daha önce ışık mikroskobuyla varlığı saptanmasına karşın elektronprotein yağ ve az miktarda karbonhidrat moleküllerinden (özellikle memelilerde) meydana gelmiştir. Hücre zarının yapısı hakkında ilk bilimsel model Danielli ve Dawson tarafıdan ortaya konmuş ve birçok biyolog tarafından uzunca bir süre benimsenmiştir. Bu modele göre hücre zarının ortasında bir fosfolipit (60 A° kalınlığında) ve bunun her iki tarafında da birer protein tabakası (30 A° kalınlıklarında) bulunur. Buna "Zar Birimi" denir.
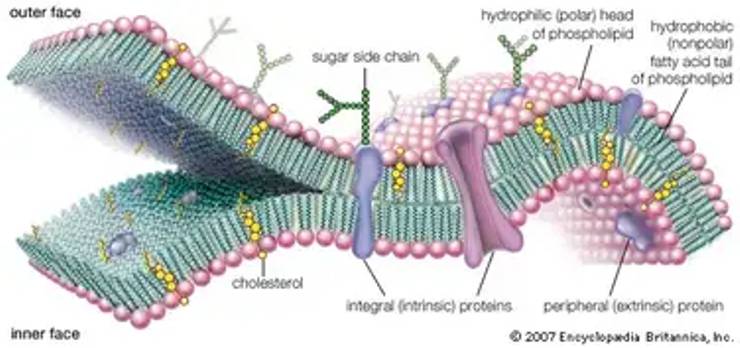
Danielli ve Dawson modelinin hücrenin işlevsel bir parçası olan hücre zarının işleyişini tam olarak açıklayamaması bu konuda yeni modellerin geliştirilmesine neden olmuştur öyleki hücre zarının iki tarafında protein ortada fosfolipit tabakasından ibaret bir yapı olmadığı; bir lipit denizinde yüzen proteinden ve glikoproteinlerden yapılmış almaç denen özel bölgelerle dışarıya açılan bir "Mozaik Zar Modeli"nden oluştuğu anlaşılmıştır. Mozaik Zar Modeli 1966 yılında Singer ve Lenard tarafından ortaya çıkarıldı ancak 1972 yılında ayrıntılı olarak yayınlandı. Bu zar modeli ya da birimi tüm hücrelerin dış zarında ve içteki organellerinin zarlı kısımlarında (mitokondri çeperi golgi endoplazmik retikulum çekirdek zan gibi) benzerdir.
Zarın yapısındaki lipitler çoğunluk fosfolipitlerdir ve zarın orta kısmında iki tabakalı olarak bulunur. Bir tabakadaki fosfolipidin suda erimez lipofil (apolar) kutbu (yağ asitlerini taşıyan polarize olmamış kutbu) öbür tabakadaki fosfolipidin lipofil kutbuna dönüktür. Dolayısıyla ışınsal bir şekilde lipofil kutuplar karşıkarşıya gelmiştir. Suda eriyen hidrofil (polar) kutupları ise dışa dönüktür. Bu tabakalar polipeptitterden meydana gelmiş bloklarla ya da adacıklarla kesilmiştir. Bu haliyle hücre zarı içinde proteinlerden yapılmış adalar taşıyan bir lipit denizi gibi görünür.
Hayvansal hücrelerin dış yüzü hücre zarında bulunan nöraminik asidin iyonize olmuş karboksil grubundan dolayı eksi yüklüdür. Nöramin nöraminidaz enzimi ile zardan koparılırsa eksi yükün büyük bir kısmı yitirilir.
Zar proteinleri yerleşim durumlarına göre iki kısma ayrılır. Bir grup protein yağ tabakasının her iki yüzündedir. Bunlara "Ekstrinsik Proteinler" denir. Bir kısmı da yağ tabakasının içine gömülmüştür; dış kısımları yağ tabakasının iç ya da dış yüzüne açılabilir. Bunlara da "l n t r i n s i k Proteinler" denir. intrinsik proteinlerden rodopsin retinanın çomakçıklarındaki taraklarda bulunur. Karanlıkta 1 /3'ü oranında. aydınlıkta ise 1 /2'si oranında zar içine gömülür. Ekstrinsik proteinler sulu ortamla temas halinde oldukları için hidrofilik amino asitleri intrinsik proteinler ise bir tarafları ile yağ tabakasına gömülü oldukları için bu kısımlarında hidrofobik amino asitleri diğer tarafları sulu ortamla temasta olduğu için de o taraflarında hidrofilik amino asitleri taşırlar.
Memeli hücrelerinde özellikle alyuvarlarda intrinsik proteinlere bağlı olarak karbonhidratlar bulunmuştur. Karbonhidratlar hücre zarında glikoproteinler ve glikolipitler halinde bulunurlar ve zar yüzeyinin türlere hatta hücre gruplanna ilişkin özgüllüğünü sağlarlar. Organellerin zarında karbonhidrat bulunamamıştır. Hücre yüzeyinde ince bir film halinde bulunan glikoproteinler hücreye antijenvirüs almacı olarak da kullanılırlar. Alyuvarlardaki mukopolisakkaritler antijen özelliğinin yanısıra kan gruplannın oluşumunu da sağlar. Bu karbonhidrat gruplarının bozulması (kanserleşme) ya da bir çeşit aşınması yani hücre zarının ketleşmesi yaşlanmaya yol açar. özellikle kanserleşmede hücre yüzeyi daha fazla eksi elektrik yüklü olur.
Çok hücrelilerde hücrelerin birbirine teması bazı bilgilerin aktarılmasına örneğin hücre bölünmesinin durdurulmasına "Kontak inhibisyon" morfo-genetik hücre hareketlerinin meydana gelmesine büyümenin düzenlenmesine neden olur. Kontak inhibisyona güzel bir örnek de plazmodyumun (sıtma etkeni) eritrositleri tanımasıdır. Bu tanımayı glikokalikslerle yapar. Diğer birçok hücre paraziti aynı yöntemi kullanır.
Hücrenin iç ortamını dış ortamdan ayıran ve her iki ortam arasındaki madde alışverişini düzenleyen hücre zarının yapısı büyük bir olasılıkla sabit değildir. Yağ ve protein molekülleri belirli sınırlar içinde hareket eder. Bu hareket içe ve dışa doğru olmaktan ziyade yanlara doğrudur. Hücre zarının yapısal değişimi taşıdığı doymuş ve doymamış yağ moleküllerinin miktarına bağlıdır. Zar genellikle vücut sıcaklığında akıcı olan doymamış yağ moleküllerini içerir. Zar yüzeyinde mozaik şeklinde bulunan protein ve glikoprotein adacıkları etrafını çeviren bir sıralı yağ molekülleri ile sıkıca bağlanmıştır (bu ikisinin arasında hareket meydana gelmez). Fakat diğer yağ molekülleriyle bağlantısı gevşektir.
Hücre zarının biyolojik etkinliğini değiştiren birçok madde örneğin karsinojen (kanserleşmeye neden olurlar) maddeler bazı hastalıkların ortaya çıkmasına neden olmaktadır.
Hücre zarının enine kesitlerinde boyları 75 A0 kadar olabilen bazı kanalların dış yüzeyden iç yüzeye kadar uzandığı saptanmıştır. Elektron mikroskobuyla yapılan son çalışmalarda hücre zarının Golgi aygıtının bir ürünü olduğu saptanmıştır. Golgi aygıtından kesecikler şeklinde sürekli meydana gelen zar akımı hücre zarının kısmi onarımında ve hücre bölünmesinden sonra hücre zarının büyümesinde kullanılır. Hücre zarında çekirdek zarında bulunan porlar bulunmaz. Hücreye giren besinleri ve hücreden çıkan artık maddeleri; zarın geçirgenliği üç tabakalı moleküler dizilişi ve özellikle proteinden oluşmuş almaç (reseptör) kısımları saptamakla beraber elektriksel yükün de bu giriş-çıkışta büyük bir önemi olduğu varsayılmaktadır. Hücre içi ile dış ortam arasındaki elektrik potansiyel farkı (m V düzeyinde) bazı maddelerin içeriye ve dışarıya pompalanmasını kolaylaştırır.
Bir amip ya da silli hayvan yaralanırsa; bu yara yeni bir zarla hemen kapatılır. Bu yeni zara plazmalemma denir. Plazmalemmayı hücre arasına salgılanan maddelerle ya da bir çeşit hücre iskeletini oluşturan hücre dışındaki daha katı selüloz (bitkilerde) ya da mukopolisakkarit ve albuminoid yapılarla karıştırmamak gerekir.
Hücre yoğunluğu az olan bir sıvı içerisine (hipotonik) konursa şişer ve sonunda patlar buna "Hemoliz" (genellikle alyuvarlarda hemin hücre dışına çıkmasında kullanılır); yoğunluğu fazla bir sıvı içerisine konursa su kaybederek büzülür hücre zarı bitkilerde selüloz duvardan ayrılır ve sonunda yine patlar buna da "Plazmoliz" denir.
Hücre zarı ya da hücre membranı, hücrenin dış kısmında bulunan, molekülleri özelliklerine göre hücre içine alan veya dışarı bırakan katmandır.
Hücre Zarı
Hücre zarını ayırarak doğrudan analizlerden önce hücre zarının moleküler yapısı hakkındaki kuramlar, dolaylı kanıtlara dayanır. Yağda eriyen maddeler hücre zarından kolayca geçebildiği için, Overton (1902), hücre zarının ince bir lipit tabakasından yapıldığını ileri sürmüştür. Gorter ve Grendel (1902), hücre zarının iki lipit molekülü kalınlığında bir tabaka (bilayer) olduğunu ileri sürmüşlerdir. Geçirgenlik, yüzey gerilimi elektrik ve kimyasal özelliklerini göz önünde bulundurarak, Danielli ve Davson 1935'de hücre zarının simetrik zar modelini teklif etmişlerdir. Bu modele göre, zarın yapısında tek tabakalı iki protein yaprağı arasında lipit molekülleri vardır. Lipit moleküllerinin polar uçları (hidrofilik kısımları) dışa doğrudur ve protein tabakalarıyla örtülüdür. Moleküler yapıyla ilgili ikinci model, Robertson (1959) tarafından teklif edilen asimetrik zar modelidir. Asimetrik zar modelinde, ortada iki molekül kalınlığında lipit tabakası, iki tarafında da tek molekül kalınlığında protein tabakası vardır.
İki model de birbirine benzemekle, arasındaki fark; birinci simetrik modelde ortadaki lipit molekül sırasının veya tabakasının kalınlığı belli değildir. Yani, iki veya daha fazla lipit molekül sırasının bulunup bulunmadığını gösteren hiçbir kanıt yoktur. Oysa, asimetrik modelde ortadaki lipit moleküllerinin sayısı sadece ikidir. İki model arasında ikinci önemli fark, lipit tabakasının iki yanındaki protein tabakalarının simetrik modele simetrik, asimetrik modele ise, kendisine eklenen yeni elementlerden dolayı sitoplazma tarafındaki protein tabakasının dıştaki protein tabakasından belli kimyasal farklar göstermesi, yani asimetrik oluşudur. Daha sonraları ortaya çıkan teori ise, Danielli-Davson'un modelidir. Danielli-Davson'a göre, lipit moleküllerinin polar, hidrofilik uçlarının koyu bölgeleri şekillendirdiği, polar olmayan, hidrofobik yağ asidi zincirlerinin açık renk bölgeleri şekillendirdiği düşünülmektedir.
Bu modellerde hücre zarı, fosfolipit elementlerin kimyasal özelliğinden dolayı iki tabakalı görülür. Bu üç tabakalı yapı, plazma zarı dışında hücrenin sitoplazmada bulunan tüm zarlı yapılarında da görülmektedir. Danielli-Davson ve Robertson modelleri, hücre zarının elektriksel ve pasif geçirgenlik özelliklerini açıklamak yeterlidir. Bununla beraber, zardaki protein elemanlarının aktif taşınmayı nasıl gerçekleştirdiğini anlamak bu modelle zordur. Danielli ve Davson modelinin, hücre zarının işleyişini tam olarak ortaya koyamamasından dolayı, yeni hücre zarı modelleri geliştirilmiştir. 1972 yılında Singer ve Nicolson tarafından hücre zarının tüm özelliklerin açıklayan bir model ileri sürülmüştür. Böylece, mozaik zar modeli ya da akışkan-mozaik zar modeli 1966 yılında Singer ve Lenard tarafından ortaya atılmasına rağmen, 1972'de yayınlanmıştır. Bu modelde fosfolipit tabakaları daha önceki modellerdekine benzer şekilde hidrofilik başları zarın yüzeyine doğru, hidrofobik kuyrukları ise, içe doğru sıralanır. Asıl farklılık proteinlerin dizilişinde görürlür.
Bu modelde proteinler zarın hem iç, hem dış yüzeyinde mozaik şekilde dağılırlar ve devamlı bir tabaka meydana getirmezler. Hücre zarında bulunan zar proteinleri; bu modelde yağ tabakasının her iki yüzünde olan ekstrinsik proteinler, yağ tabakasının içine gömülmüş olanlar ise; intrinsik proteinler olarak kabul edilmiştir. Bir lipit denizinde yüzen, protein ve glikoproteinlerden yapılmış, almaç denilen özel bölgelerle dışarıya açılan bir model olarak mozaik zar modeli günümüzde de geçerliliğini korumaktadır.
Moleküler yapısı
Hücre zarının moleküler yapısı hakkında bilgiler, kimyasal analizlerden, yaşayan hücrelerin yüzey gerilimi, elektrik ve geçirgenlik özellikleri gibi farklı fizikokimyasal özelliklerinden, antijenik özelliklerinden, polarizasyon, X-ışını difraksiyonu ve elektron mikroskobu gözlemlerinden elde edilmiştir.
Elektron mikroskobuyla yapılan çalışmalarda, hücre zarlarının ortada açık renk bir tabakayla ayrılan iki koyu tabaka olmak üzere üç tabakalı bir yapı olduğu gösterilmiştir. Bu yapı, Danielli-Davson ve Robertson tarafından bildirilen modellere uygundur. Bu modelde, fosfolipit tabakalarının hidrofilik başları zarın yüzeyine doğru, hidrofobik kuyrukları ise, içe doğru sıralanır, proteinler zarın hem iç, hem dış yüzeyinde mozaik şekilde dağılırlar ve devamlı bir tabaka meydana getirmezler. Akışkan zar modelinde zar hareketsiz değildir, birbirine zayıf bağlarla bağlı olan bireysel lipit molekülleri lateral olarak hareket edebilirler.
Buna göre, herhangi bir molekül belli bir zamanda belli bir pozisyonda bulunurken, birkaç saat sonra tamamen farklı bir pozisyonda bulunabilir. Lipitlerin hareketi en fazla kolesterol içermeyen zarlarda görülür. Proteinler de belli sınırlar içinde lateral olarak hareket edebilirler. Fakat proteinlerin hareketi lipitlerinkinden daha azdır. Lipit yapraklarını baştan başa kat eden, iki yüze de açılan zar proteinleri şekillerine göre kabaca ikiye ayrılır. Bunlar; çubuk şeklinde ve küre şeklinde zar proteinleridir. Bu proteinler hücre dışı moleküller olarak reseptör görevinde ve bağışıklık sisteminde yabancı maddeleri tanımada rol oynarlar.
Hücre zarında bulunan zar proteinleri; yağ tabakasının her iki yüzünde olan ekstrinsik proteinler, yağ tabakasının içine gömülmüş olanlar ise; intrinsik proteinler olarak bilinir. İntrinsik proteinler, karanlıkta 1/3'ü oranında, aydınlıkta ise, ½'si oranında zar içine gömülüdür. Ekstrinsik proteinler sulu ortamla temas halinde bulundukları için, hidrofilik aminoasitleri, intrinsik proteinler ise bir tarafları yağ tabakasına gömülü olduğu için bu kısımlarına hidrofobik aminoasitleri, sulu ortamla temas halinde olan diğer taraflarında ise, hidrofilik aminoasitleri taşırlar.
Hücre zarında, çekirdek zarında bulunan porlar bulunmaz. Hücreye giren besinleri ve hücreden çıkan atık maddeleri; zar geçirgenliği, üç tabakalı moleküler diziliş ve özellikle proteinden oluşmuş almaçlar (reseptör) ile elektriksel yükün de önemi olduğu düşünülmektedir. Bir hücre zarından zardan her türlü madde geçebiliyorsa bunlara geçirgen (permeabl), hiçbir maddeyi geçirmiyorsa geçirgen olmayan (impermeabl) ya da geçirimsiz, bazılarını geçiriyor ve bazılarını geçirmiyorsa da seçici geçirgen (semipermeabl) hücre zarı denir.
Tekhücreli bir canlıdaki hücre zarında bir yara oluşursa, bu yara yeni bir zarla hemen kapatılır, bu yeni zara plazmalemma denir.
Yan yana duran iki hücrenin sitoplazma zarları arasında 150-200 Å (angstrom) genişliğinde hücrelerarası bir alan vardır. Bu alan, hücreleri birbirine yapıştıran bir madde ile doludur. Hücre zarı girintili çıkıntılıdır. Bu yapı hücreler arasında adezyonu ve aynı zamanda hücreler arasındaki dokunma yüzeyini artırır.
Plazma zarının sitoplazmaya bakan yüzünde zar elemanları bulundukları noktalara demirleyen sitoiskelet elemanları yer alır. Sitoiskeleti oluşturan elemanlar şunlardır:
* Mikrofilamenteler
* Kalın filamentler
* Mikrotübüller
Mikrofilamentler ve mikrotübüller reseptörlerin kontrolünde iş görürler. Mikrofilamentler kasılarak reseptörlerin hareketini idare ederler. Mikrotübüller ise, demirleme elemanlarıdır. Reseptörleri tutarlar veya serbest bırakırlar.
Hücre zarının kimyasal yapısı
* Zar lipitleri
* Fosfolipit
* Glikolipit
* Sterol
* Zar proteinleri
* Zar karbonhidratları bulunur.
Hücre zarının morfolojisi
Elektron mikroskobunda hücre zarı oldukça basit yapıda görülür. İki koyu ve bir açık renk olmak üzere üç tabakalı görülen yapıya, üç tabakalı veya unit zar, birim zar, denir. Bu tabakaların kalınlığı 75-100 Å arasında değişir. Lipitlerin hidrofobik kuyrukları açık renk görülür. Lipitlerin hidrofilik uçlarıyla proteinler birlikte koyu çizgiler oluşturur.
1- Glikokaliks
2- Hücre zarı farklılaşmaları (Desmosomlar)(Desmosomes, Cell Junctions, Cell Attachments)
* Aralarında farklılaştıkları hücrelerin benzer veya farklı oluşuna göre:
Otodesmosomlar
Homodesmosomlar
Heterodesmosomlar (Hemidesmosomlar)
* Hücre yüzeylerinde dağılışlarına göre:
Makular desmosomlar
Zonular desmosomlar
* Simetri durumuna göre:
Simetrik desmosomlar
Simetrik olmayan desmosomlar
* İki hücre yüzeyinin elemanlarının desmosom yapısına katılamalarına göre:
Sinsisyal desmosomlar
Basit desmosomlar
Gelişmiş desmosomlar
Hücre zarının görevleri
Hücre zarı, oldukça karmaşık ve devingen yapısıyla, hücre canlılığının çok önemli bir bileşenidir. Hücre canlılığının ve özgün hücre işlevlerinin sürekliliğini mümkün kılan çok önemli bazı fonksiyonları yerine getirir ki, bunları şöyle sıralamak mümkündür:
* Hücre içi ortamın özgün bileşimini hücre dışı ortamdan ayırmak,
* Hücre içi ile hücre dışı ortamlar arasında seçici bir şekilde madde alışverişini sağlayarak hücrenin atıklarını hücre dışı ortama vermek, hücre dışından hücreye gerekli maddeleri almak ve hücre içi ortamın özgün yapısını korumaya yardımcı olmak,
* Komşu hücrelerle iletişimi ve madde alışverişini sağlamak,
* Hücreyi dış ortamdan ayırır.
* Hücreye şekil verir.
* Madde giriş-çıkşını düzenler.
* Canlı yapıdadır.
* Kalınlığı 6-10 nm'dir.
* Protein, yağ ve karbonhidratlardan oluşur.
* Aktif taşıma olayını düzenler.
* Hücrenin beslenmesine yardımcı olur.
* Komşu ve yabancı hücreyi bulur.
* Hücreyi alınacak hormonları tanır.
* Hücrenin yıpranan kısmını onarır.
* Metabolizma atıklarının dışarı atılmasını sağlayarak iç ortamı düzenler.
* Prokaryot hücreye sahip canlılarda zardaki solunum enzimleri sayesinde enerji üretimi sağlanır.